

1. Две проблемы описания живых систем в формулировке Э. Шредингера
В своей широко известной книге 1943-44 г.г. [1] «Что такое жизнь? С точки зрения физика» Э. Шредингер на первое место выводит главное для ответа на вопрос в её заголовке – нужна на языке законов и методов физики и химии теория, обобщающая огромное количество накопленных в биологии фактов. Как основу такой будущей теории он принимает два вопроса, возможным путям ответа на которые посвящена вся его книга.
Первый из них о том, что сегодня называют генетической основой жизни. Прочитав книгу Шредингера, Ф. Крик бросил физику и занялся биологией. В результате всего через 10 лет после этой книги месте с Дж. Уотсоном и Р. Франклин была установлена структура ДНК.
За прошедшие с тех пор полстолетия в исследованиях ДНК накоплен большой объём фактов. До окончательности в их сборе ещё далеко. О них и их истории написано столько, что повторять это здесь в порядке обзора хотя бы кратко слишком увеличит объём статьи.
Второй поставленный Э. Шредингером вопрос относится к совместимости существования и эволюции жизни со вторым началом термодинамики. Самого этого термина Шредингер не использует, заменяя его вопросом – жизнь является «порядком из порядка» или «порядком из беспорядка»? (см. примеч. 1). Ответа на этот вопрос он найти не смог. Более того, последнюю главу своей книги он начинает словами «...я хочу показать, что всё известное нам о структуре живой материи заставляет ожидать, что деятельность живого организма нельзя свести к проявлению обычных законов физики». Отсутствие ответа на этот вопрос привело сегодня генетику на намного более высоком уровне знаний к тому же состоянию, с которого Шредингер начал в 1943 г. свою книгу – нет опережающей физической теории, которая создала бы стержень для систематизации и использования биологических фактов.
Этот парадокс устранён 50 лет спустя после книги Шредингера в работах [2] – [8]. Из них следует, что основ существующей физики достаточно для описания жизни, её возникновения и развития, а также разума и сознания человека, но необходимо уточнение понятия об информации как физической переменной и введение в физику закона иерархического синтеза действия-энтропии-информации (см. примеч. 2). Об этом новом законе и его следствиях для биологии и дарвинизма рассказывается в этой статье.
--------------
1. Второе начало термодинамики является одной из самых первичных аксиом современной науки. Однако вопреки законам об аксиомах существует более 20 определений второго начала термодинамики (подробнее см. [2] глава VI, параграфы 3, 4). Можно догадываться, что именно это заставило Шредингера прибегнуть к замене, так как речь у него идёт о неназванном одном из таких определений.
2. Кстати, на основе этого же закона в [6] устраняется общеизвестный парадокс отсутствия логических связей между классической и квантовой механикой.
2. Определение информации как физической переменной
Понятие об информации возникло примерно в начале Второй мировой войны. Развивалось оно в значительной степени под влиянием практических задач передачи обычных и шифрованных сообщений. Поэтому начальная доля закрытых научных результатов не была малой. В России к этому добавилось, что к «продажным девкам» с соответствующим цензурированием была отнесена не только генетика, но и кибернетика. Выход в свет в 1958 г. научно-популярной книги И.А. Полетаева «Сигнал. О некоторых понятиях кибернетики» [9] был полукрамольным событием. Последствия цензурирования в отношении генетики и кибернетики по сей день мешают (особенно в биологии и генетике) пониманию всего, что связано с информацией и ролью в ней энтропии.
Истоком современной теории информации для средств связи человека и компьютеров являются работы К. Шеннона. Он определил информацию как устранённую неопределённость в достижении цели. Такое определение вошло во все учебники теории информации, например [10], [11]. Слово – цель – при этом в связи с очевидностью опускается. Формальное математическое описание неопределённости у него совпало с принятым в физике для энтропии.
В современной физике стало рутинным приёмом правильные интуитивные находки вводить в описание природы, не объясняя первичные связи с известным, которыми руководствовались авторы. Наиболее значительный пример такого дал П. Дирак (пояснения см. [12], глава IV, параграф «Забытое или “ошибки”»). Подобное содержат основы Стандартной модели элементарных частиц в физике. Не стал объяснять и Клод Шеннон связи своего термина энтропия в теории информации с энтропией в физике. Это начал Л. Сцилард, но Вторая мировая война разбросала учёных по всему свету и он сделал меньше того, что мог бы. На успехах теории информации в задачах техники это не сказалось.
Левая колонка на рис.1 вводит ключевые слова, с помощь которых каждый может вспомнить или прочесть заново по учебникам известное об информации в теории связи, а также см. [2], глава I, параграф 3 «Что такое – информация?».
 Рис.1
Рис.1
В физике определение понятия информации пошло по пути аналогий с решениями для техники связи. Общепринятое сегодня её определение в физике связано с именем выдающегося физика Л. Бриллюэна (см. примеч. 3). Обычно его поясняют на примере энтропии газа. Состояние газа как системы составляющих его молекул задано макроскопическими параметрами в виде объёма, давления, температуры, химического состава и др., например, в каких-то задачах в это могут входить электромагнитные переменные и параметры. Микроскопически этот же газ описывается состояниями каждого из атомов в нём. Число таких разных микросостояний, соответствующих одному и тому же макросостоянию, огромно. Выражается оно логарифмической функцией, которую называют – энтропия. Так как числомикросостояний стоит в ней под знаком логарифма, то абсолютные величины энтропии в природе в большинстве случаев с запасом укладываются в сотню. Но это не меняет ситуации – человек не в состоянии описать физическую систему с помощью воспроизведения каждого микросостояния. Тем не менее, информация в физике определена с участием энтропии в виде: энтропия системы в данном макросостоянии есть количество информации, недостающее до её полного описания. Но ведь природе безразлично, чего недостаёт человеку для её описания. Устранённая неопределённость – это и есть информация. В технических задачах человека или в природе – безразлично. Что такое устранение неопределённости в природе? Ответ на этот вопрос дан в работах [2] – [8]. Поясню его здесь. Объём газа, камень на дороге, живая клетка, галактики в космосе существуют как макроскопические объекты. Самим фактом своего существования они устранили неопределённость микросостояний составляющих их элементов . Отсюда можно сформулировать определение: информация как физическая переменная есть сама величина энтропии микроэлементов, составляющих макроскопический физический объект (см. примеч. 4) Введенное в [2] – [8] такое определение информации как физической переменной даёт твёрдую основу для описания на языке математики жизни и разума в терминах физики и химии, то есть создаёт основу, чтобы снять вопросы, на которые не было ответа у Шредингера в его книге [1] (см. примеч. 5). Подробные объяснения к правой колонке на рис.1 содержатся в [2] – [8] и ниже в этой статье.
Обстоятельства сложились так, что в октябре 2000 г. Л.А. Блюменфельд снимал последние вопросы редактора своей книги [14], написанной первично на английском. В ней содержится итог для биологической физики того, что возникло в науке на основе определения Л. Бриллюэна для информации в физических системах. И одновременно с этим он написал отзыв на книгу [2], который был опубликован в журнале «Биофизика» РАН [15]. Этим совпадением он отметил важность новых решений в биологической физики, которые вводит монография [2] и предыдущие работы, на которых она основана.
-----------------
3. Его имя входит в названия семи крупных результатов в физике.
4. Энтропия является положительно определённой переменной. Она аддитивна, то есть нуль её отсчёта, если она вводится как иерархическая переменная действие-энтропия-информация, может быть выбран на любой ступени иерархии. Это разрешает обозначать при необходимости слагаемые до этого нуля отрицательным знаком, но такое не имеет никакого отношения к негэнтропии Бриллюэна.
5. В [13] микробиолог Г.А. Заварзин пишет, цитирую: «Я могу сказать совершенно однозначно, что как только я слышу слово «информация», я немедленно выключаю свои умственные способности. Спекуляции на слове «информация» настолько распространены, что ... (многоточие в оригинале)». В силу объяснённого выше могу в этом понять Г.А. Заварзина – результаты, полученные на основе исходно некорректного определения информации (каковые и составляют реальность литературы) рациональный человек принимает с трудом. Но хотят этого биологи или нет, после того как в [2] – [8] и здесь эта некорректность исправлена, дальше игнорирование в биологии и генетике понятия об информации в строгом виде [2] – [8] становится тормозом для этих областей науки.
3. Порядок (информация) в природе самопроизвольно возникает из беспорядка
Со времён Лапласа детерминизм в природе определяется аналогией с траекторией материальной точки классической механики. Существуют уравнения механики, которые позволяют вычислить её с любой заданной точностью. Фактически такая траектория в XIX веке стала эталоном того, что называют – порядок в природе.
Однако в ХХ веке выяснилось, что если движение одиночной материальной точки происходит в замкнутом объёме, у которого хотя бы одна стенка выпуклая вовнутрь или если в неограниченном объёме таких материальных точек много и они взаимодействуют между собой, то эталон детерминизма и порядка превращается в одну из самых быстро нарастающих неустойчивостей в природе, то есть становится синонимом непредсказуемости, беспорядка. Подробнее об этом см. [2], [7].
Ещё Л. Больцман обратил внимание, что для системы из многих взаимодействующих элементов положение и острота максимума их статистической характеристики – энтропии – определены с огромной точностью (если этих элементов много, например, как в реальности для газа). Это оставалось недостаточно понятым как во времена книги Шредингера, так для многих и сейчас. В работах [2] – [8] впервые энтропия введена как иерархическая переменная действие-энтропия-информация. Иерархия возникает из-за смены признаков, относительно которых определяется энтропия реальных систем. Здесь действие есть переменная классической механики. Тогда известный принцип наименьшего действия Мопертюи определяет классическую траекторию материальной точки механики как экстремум энтропии, каковой становится в механике действие. Тем самым классические лапласовский детерминизм и порядок при их выражении траекторией единственной изолированной материальной точки совпадают с определением детерминизма и порядка в виде положения и остроты экстремума энтропии (действия-информации). В силу объяснённого, альтернатива Э. Шредингера в [1] – «порядок из порядка» или «порядок из беспорядка» – теряет свою остроту. В природе «порядок» закономерно возникает из «беспорядка» в форме экстремума описывающей систему энтропии (действия-информации). При этом закон иерархического синтеза действия-энтропии-информации вводит в «порядок из беспорядка» важную особенность. Поясню её.
Понятия – объект (включая в это слово для краткости и процесс) – означает, что нечто, существующее в природе статически или динамически, воспроизводимо, то есть в одинаковых условиях будет тождественным, как, например, химические элементы во всей Вселенной. Этим факт существования объектов природы становится синонимом их устойчивости в строгих терминах физики и её языка математики. Энтропия является функцией Ляпунова в задачах физики, анализ которой даёт ответ об устойчивости. Роль функции Ляпунова в некоторых задачах может выполнять энергия. Для определения устойчивости существуют и другие критерии, не использующие функции Ляпунова. Это не меняет главного – устойчивость неразрывно связана с понятием информации.
Информация в природе (однородно с теорией связи) определена как устранённая неопределённость (см. параграф выше). В природе устраняет неопределённость состояний элементов, образующих её объект (микросостояний), сам факт его макроскопического существования. Поэтому объекты природы возникают и существуют на основе информации о них как их первичной сущности. В теории информации для средств связи существует, начиная с середины ХХ века, с книги [16] Г. Кастлера, понятие – синтез информации. В работах [2] – [8] оно дополнено и распространено на физико-химические объекты природы.
Микросостояния физических систем случайны, но не обязательно эти случайности независимые. Информация как физическая переменная синтезируется на их основе. Не важно, какие функции распределения имеют эти случайности, но в природе редко встречаются варианты, когда они могут считаться независимыми.
Случайности есть первая составляющая синтеза в природе информации как физической переменной, см. схему на рис.2.
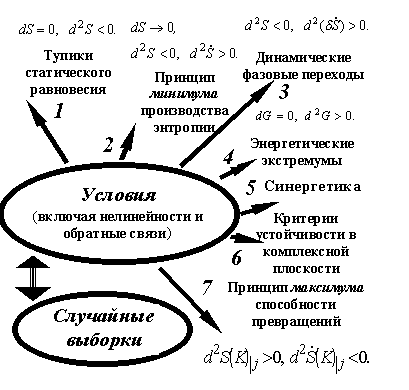 Рис.2
Рис.2
Поясню о случайностях более наглядно. Выпадение очков игральной кости является классическим примером независимых случайностей. При этом многие напрочь забывают, что это сугубо частный случай реализации случайностей. Реально они всегда ограничены условиями. Простейший пример: при игре в кости никогда для одной кости не выпадет число очков больше 6. Кроме того, в терминах аналогии с игрой в кости природа преимущественно пользуется «мошенническими костями», то есть случайности (как основа для работы естественного отбора) в ней редко являются строго независимыми, равновероятными. Однако именно в таком частном смысле случайности понимаются в дарвинизме и его уточнении с помощью синтетической теории эволюции. В результате такое огрубление становится основой большинства противоречий и парадоксов дарвинизма.
На рис.2 показаны классы процессов и критериев их устойчивости, которые в природе в составе синтеза информации (действия-энтропии) превращают «беспорядок» в «порядок» [2] – [8].
Первый из них (1 на рис.2) описывает идеализации объектов как замкнутых систем. Понятие замкнутых (закрытых систем) означает равновесие на их границах. Ими могут быть разные сосуды или непосредственно поверхность объектов. Критерий1, например, задаёт равновесное сохранение ДНК в спорах бактерий.
Максимум энтропии, описывающей поведение элементов, составляющих замкнутую систему, является условием её существования. Как отмечалось выше, сама энтропия (действие-информация) объекта в виде замкнутой системы есть содержащееся в нём количество информации как физической переменной.
Подвод к системе, например, энергии делает её открытой. Она самопроизвольно стремится вернуться к потерянному равновесию. Реально в этом участвуют диссипативные процессы, то есть производство энтропии и его возмущения2, 3 на рис.2). Необходимо подчеркнуть, что в случаях 2 и 3 энтропия растёт. Ограничение накладывается только на темп роста – при прочих равных условиях он должен быть минимальным. При критериях динамической устойчивости2, 3 на путях возврата формируются динамические равновесия. Их И. Пригожин назвал «возникающим» в отличии от «существующего» в виде объектов типа 1 на рис.2. Диссипативной самоорганизации посвящен огромный объём литературы. Здесь надо подчеркнуть, что устойчивость (то есть реальность «возникающего» согласно критериям 2, 3 на рис. 2 для производства энтропии и его возмущений) существенна для ряда объектов и процессов в живой природе. Результаты применения к ним этих критериев известны, значительны, но здесь делать их обзор нецелесообразно.
И для 1, и для 2, 3 на рис.2 энтропия может пониматься в обычном смысле теплотехники или физической химии. Однако в общем виде для схемы на рис.2 энтропия остаётся действием-энтропией-информацией. Следствия этого для дарвинизма введены в [2]. Они исключительно существенны и объяснены в конце этого параграфа.
Энергия в роли функции Ляпунова приводит к классу объектов, устойчивость которых зависит от экстремумов свободной энергии как одного из термодинамических потенциалов (4 на рис.2). Наиболее известными из них являются свободная энергия Гиббса и Гельмгольца. Экстремумы свободной энергии лежат в основе физической химии как области науки. Если в задачах участвуют ещё электрические и магнитные переменные, то число термодинамических потенциалов и соответственно их экстремумов, удобных в частных задачах, увеличивается. Устоявшихся названий для них нет. В таких случаях методы физической химии оказываются результативными при описании равновесий в химических реакциях с участием электромагнитных полей. Фактически в терминологии Э. Шредингера критерии типа 4 на рис.2 можно было бы считать ответственными за превращения «порядка в порядок». Но как отметил в [17] Л.А. Блюменфельд, экстремумы свободной энергии не могут дать идеальной упорядоченности (например, понимаемой как симметрии кристаллов или макромолекул). Какие бы формы энергии ни входили в выражение для изменения полной энергии dU, уравнение для приращений dF свободной энергии
dF = dU - T dS
включает в себя в последнем члене энтропию (T – температура, S – энтропия). Поскольку рассматриваются процессы физической химии, то энтропия здесь имеет обычный смысл. Макроскопически участие энтропии нарушает идеальный порядок, например, кристаллов. В них, как минимум, появляется равновесная концентрация дефектов кристаллической решётки. То есть (в отличие от общепринятого толкования) критерии устойчивости объектов на основе экстремумов свободной энергии часто вводят в системы беспорядок. Поэтому иногда необходимо перейти от чисто энергетических критериев при описании синтеза информации к критериям устойчивости в общем виде, когда не нужны априорные предположения о соотношении энергетических и энтропийных критериев устойчивости. Это сделано в [2] путём введения понятия об энтропии (действии-информации) на математическом языке функций комплексного переменного. Тогда критерии устойчивости можно формулировать способами, известными в теории автоматического регулирования (6 на рис.2).
В задачах развития процессов во времени может использоваться анализ систем линейных и нелинейных дифференциальных уравнений первого порядка. Обычно такое называют динамическим описанием систем. Его относят к созданной Г. Хакеном области науки, которую он назвал синергетика. В широком смысле к синергетике относят задачи развития и устойчивости структур, которые зависят от многих факторов и включают в себя нелинейности. Эта область анализа устойчивости обозначена 5 на рис.2. При динамическом описании систем на первый план выходят точки бифуркации решений (термин веденный А. Пуанкаре). Это точки, в которых решение в результате сколь угодно малого случайного воздействия может пойти по одному из двух путей. Система хаотизируется. Такое часто трактуют как непредсказуемость результата. На самом деле это только предупреждение, что в данной задаче систему эффективнее описывать статистическими, а не динамическими, методами. Непредсказуемость в точках бифуркации индивидуальных уравнений совместима с однозначностью результата, например, в терминах экстремумов энтропии и её производства для системы в целом.
Критерии 1 – 6 на рис.2 создают новое при возвратах систем (в частности живых) к равновесиям (преимущественно динамическим) после или в процессе подвода к открытой системе энергии и веществ. Состояния системы при этом возникают новые, но в своей основе это оригинальные остановки на пути восстановления прошлого равновесия. Тогда в новой форме на первый план выходит парадокс ещё У. Томсона (лорда Кельвина) «тепловой смерти Вселенной», так как новые объекты могут возникать только на путях к некоему глобальному или частному равновесию как пределу.
Устраняет этот парадокс критерий устойчивых потоков, сопровождающихся ростом энтропии (действия-информации) за счёт изменения признаков, относительно которых определяется энтропия (действие-информация). Он показан 7 на рис.2. Это критерий максимума способности превращений. Он описывает случаи, когда система открывает для себя класс динамических или статических равновесий, которые основаны на иерархически новых признаках для действия-энтропии-информации, то есть осуществляется переход по ступеням иерархии этого роста.
Как известно из теории информации, энтропия при дополнительных условиях меньше, чем без них. Поэтому интервал изменения энтропии (действия-информации) на каждой следующей ступени иерархии её роста меньше, чем на предыдущей. Это показано на рис.3.
 Рис.3
Рис.3
Отсюда кажущееся убеждение, что развитие (эволюция) жизни сопровождается упорядочением систем. Однако реально беспорядок (строго – действие-энтропия-информация) только растёт, но со сменой признаков, то есть иерархически. С учётом этого развитие в природе и в частности жизни можно представить схемой на рис.4.
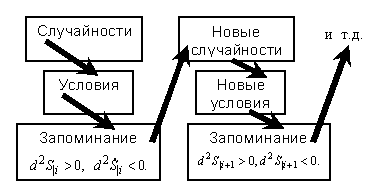 Рис.4
Рис.4
Часто забывают, что термин энергия появился в науке в то же десятилетие, что и книга Дарвина «Происхождение видов», а термин энтропия Р. Клаузиус ввел в 1865 г., то есть на 7 лет позже неё. Дарвин их не мог знать, когда писал свою книгу. Термина – информация – не существовало ещё лет сто после книги Дарвина.
Поскольку жизнь в конечном итоге сводится к законам физики и химии, то в своё время, хотя бы по этой причине, Дарвин не мог дать её исчерпывающего описания. Но его результаты сохраняются в науке уже 150 лет потому, что он правильно выбрал методологию своей работы и чётко сформулировал её основную предпосылку: «... я смотрю на все существа не как на результат отдельных актов творения, а как на непосредственных потомков немногочисленных существ, живших задолго до отложения первых пластов кембрийской системы...» (подробности см. [2], глава III, параграф 1 «Великая сила извращения чужих мыслей»). В этом параграфе впервые сформулировано главное, что определяет место дарвинизма в общей картине природы – дарвинизм и естественный отбор является эмпирической реализацией принципа И. Пригожина минимума производства энтропии с учётом иерархии смены её признаков, приводящей к действию-энтропии-информации (2, 3 на рис.2).
Повторю, что не было у Дарвина в арсенале понятий, которые возникли в физике за 150 лет после его книги. Но эмпирически Дарвин их предвосхитил. Воспользуюсь цитатой из Дарвина в [18], в которой он это объяснил: «Строение изучается не как продукт селекции, а как ограничение её» (выделение моё. А.Х.). Аргументом действия-энтропии-информации на уровне видов жизни является число состояний, выраженное в этом случае их разнообразием. Минимизация производства действия-энтропии-информации в соответствии с критериями 2, 3 на схеме рис. 2 это и есть на современном языке «ограничение её <селекции>», о котором пишет Дарвин. То есть сам Дарвин эмпирически определил область применимости своего естественного отбора теми случаями, которые сейчас получили строгое выражение в терминах физики как принцип минимума производства энтропии.
4. Жизнь – следствие единой с неживой природой иерархии развития на основе экстремумов действия-энтропии-информации и её производства
Энтропия (действие-информация) аддитивна, то есть её иерархический рост можно отсчитывать от любой ступени иерархии, приняв её нулевой. Для задач возникновения и эволюции жизни достаточно принять нулевым уровень иерархии, элементами на котором являются химические элементы таблицы Менделеева. Для их комбинаций (числа возможных состояний как аргумента действия-энтропии-информации), максимуму отвечает углерод. Это отражено вверху на рис.5. По числу соединений водород его превосходит, но у него только одна валентность, поэтому в терминах комбинаторики, используемой человеком для подсчёта величины энтропии, он с углеродом конкурировать не может.
Условие прочности химических связей выделяет элементы, которые наиболее часто встречаются в живых организмах. Они перечислены на рис.5 ближе к его середине. Этот список не абсолютен. В живых системах в меньших количествах участвуют другие химические элементы.
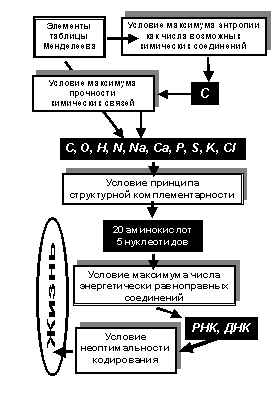 Рис.5
Рис.5
Оказалось, что среди соединений углерода есть такие, для которых справедлив (известный в биохимии) принцип структурной комлементарности – на их основе возможны цепочки до 20 последовательных практически безотходных химических реакций, в которых продукты одних используются другими. Это выделяет 20 аминокислот и 5 нуклеотидов, на которых основаны наиболее распространённые формы жизни на Земле. У термофильных бактерий возможны, но пока детально не исследованы, замены представителей этих классов соединений.
Иначе говоря, предбиологические основы жизни порождены максимумом действия-энтропии-информации в виде углерода и его химических соединений, из которых условия энергетических экстремумов (определяющих форму и свойства биомолекул) выделяют класс объектов биохимии, удовлетворяющих принципу структурной комплементарности.
Предбиологические основы жизни на планетах звёзд во Вселенной имеют единую биохимическую основу, хотя в деталях её реализации возможны варианты, сохраняющие общность принципов. Это сформулировано в [2] глава II, параграф 3 «Возникновение жизни во Вселенной – ординарное воспроизводимое явление». Важнейшая особенность предбиологических молекулярных основ жизни – возникновение как их итога нового уровня иерархии признаков «неживых» молекулярных элементов систем. Написаны о РНК и ДНК сотни книг и тысячи статей, но практически вне внимания в них остаётся важнейший факт. В химии энергетический экстремум, например свободной энергии (рис.2), критерий 4), задаёт однозначные пространственные формы молекул (зеркальные изомеры только подчёркивают это правило). В отличие от этого такой же энергетический экстремум для РНК и ДНК разрешает разные формы этих молекул как последовательностей кодонов в них. То есть однозначные энергетические экстремумы приводят к случайности форм результирующих биомолекул РНК и ДНК. Поэтому условие максимума числа энергетически равноправных разных форм молекул РНК и ДНК на физико-химическом (молекулярном) уровне иерархически выделяет жизнь в составе неживой природы, середина на рис.5.
В предбиологической эволюции важнейшее в том, что энергетические экстремумы для термодинамических потенциалов типа свободной энергии приводят к неоднозначности форм специфических молекул – РНК и ДНК, в которых разрешёны разные (не контролируемые энергией взаимодействия) комбинации последовательности кодонов. В этом состоит наблюдаемый итог предбиологической эволюции – возникновение нового признака (нового источника случайностей) как основы для роста действия-энтропии-информации. Это остаётся одним из «неживых» процессов и в конечном итоге (как и принцип структурной комплементарности) предопределено законами заполнения и взаимодействия электронных оболочек атомов. Собственно жизнь является иерархическим скачком, разрешающим разные виды синтеза информации (самоорганизации) на рис. 2 на основе новых элементов систем в виде «неживых» РНК и ДНК. Накапливаются данные экспериментов, воспроизводящих химические реакции, которые могут иметь отношение к предбиологической эволюции. Продвинуты в последнее время исследования РНК. В частности, за них присуждена Нобелевская премия 2006 г. Эндрю Файеру и Крейгу Меллоу, а в 2002 г. Большой золотой медали им. М.В. Ломоносова удостоен А.С. Спирин. Здесь обсуждать их не буду.
Надо подчеркнуть, что вся химия (а не только молекулярная биология и генетика) есть наука о «размножении» молекул. Кавычки полезны потому, что в неорганической и в обычной органической химии реакции требуют поставки извне исходных веществ и отвода их конечных продуктов. В отличие от этого на уровне, начиная от РНК, работает принцип структурной комплементарности, то есть исходные продукты и удаление конечных в большинстве обеспечено им самим. Отмечается это словами – самовоспроизведение РНК. Эти слова для РНК и ДНК имеют разный смысл, возникающий как качественное отличие в результате количественного увеличения роли случайностей для ДНК в комплексе с условиями, включающими свойства РНК.
Возникновение неоднозначной молекулы ДНК и её связь с участием РНК и принципа структурной комплементарности в синтезе информации о белковых системах (самоорганизации, включая самовоспроизведение РНК и ДНК) – это и есть жизнь во всех её формах. В её основе случайности разных пространственных последовательностей кодонов (как принято называть – генетического кода) и неоптимальность кодирования им процессов с участием белков (см. [2], глава II, параграф 9 «Отсутствие абстрактной оптимальности кодирования как особенность РНК и ДНК»). Это показано внизу на схеме рис. 5.
В силу объяснённого у прокариот нет предшественников в смысле дарвинизма, так как они возникли на основе принципа максимума способности превращений. Глобальная физико-химическая причина появления прокариот в том, что они как новая ступень иерархии роста энтропии (действия-информации) увеличивают иерархическую энтропию Земли как планеты в целом по отношению к неживому статическому пределу (см. [2], глава VI, параграф 10 «Жизнь возникает и существует потому, что увеличивает энтропию планеты сверх предела неживого равновесия»). Самая характерная особенность первичных при возникновении жизни бактерий – геологические в масштабах коры Земли последствия их метаболизма. Они рассмотрены, например, в [20], [21]. Ссылки на предыдущие работы Г.А. Заварзина см. в [2].
Дальше огромные масштабы распространения прокариот приводят к взаимодействиям между ними самими. В этом на первый план выходят недостатки эмпиричности принципа борьбы за существование как уничтожения слабейшего. В строгом смысле «борьба за существование» – это реализация принципа И. Пригожина минимума производства энтропии в применении к действию-энтропии-информации. В этом на первый план выходят математические методы, которые уже дали многие результаты. Например, среди них известные как взаимодействия «хищник-жертва». В конечном счёте такие отношения являются установлением динамического равновесия, то есть в общепринятой терминологии симбиоз (см. [2], главы II, III). Эмпиричность в дарвинизме принципа минимума производства энтропии окарикатуривается 150-летней абсолютизацией агрессивности при естественном отборе.
5. Обобщающая теория образования новых таксономических градаций жизни
Спустя полстолетия после книги Э. Шредингера [1] круг замкнулся. Успехи за это время генетики огромны. Но опять, на новом уровне количество фактов стало столь большим, что описание образования новых таксономических градаций нуждается в опережающей теории в соответствии с тем, что писал в своей книге Шредингер. Такая теория сформулирована в [2], глава IV, параграф 6 «Спонтанные и индуцированные процессы как составляющие мутагенеза». Поясню её существо.
Критерии устойчивости 1 – 7 на рис.2 позволяют детально описать синтез информации о новых таксономических градациях жизни или сохранении и видоизменении существующих. Общее для всех таксономических градаций состоит в их динамической равновесности. Процесс установления равновесий, независимо от деталей их реализаций, подчинён единому обобщающему принципу, который впервые сформулировал А. Эйнштейн как основу для повторного по отношению к М. Планку выводу закона излучения черного тела. Достаточно популярно для биологов об этом законе см. [22], глава V, параграф 6 «Повторный вывод формулы Планка Эйнштейном».
В природе равновесие возможно при равенстве скоростей процессов, направленных к нему, со скоростями деструктивных процессов, препятствующих его достижению. Случайно такое равенство практически невозможно. При описании статических и динамических равновесий на основе математического аппарата (критерии 1 – 7 рис. 2) в нём самом содержатся детали, вводящие равенство этих скоростей. Однако в биологии и генетике не всегда есть достаточно сведений, чтобы поставить задачи строго математически. Аналогичная ситуация была (и в значительной мере сохранилась) для описания акта перехода электрона между энергетическими уровнями в атоме при излучении и поглощении света атомами веществ. Причины этого лежат в основах квантовой теории. Выход из положения Эйнштейн нашёл в том, что ввел два феноменологических коэффициента. Один из них описывает частоту спонтанных переходов электрона в атоме, которая зависит от свойств этого атома и рассматриваемых энергетических уровней для начального и конечного состояния электрона. Этот коэффициент гарантирует возможность изменений в атоме, результатом которых может стать равновесие между излучением и веществом. Второй коэффициент связан с понятием об индуцированных переходах электронов, интенсивность которых зависит от плотности излучения соответствующей частоты в окрестности атома. Именно этот коэффициент обеспечивает в данных условиях возможность равенства между скоростями процессов, направленных к равновесию, и деструктивными.
Феноменологические коэффициенты спонтанных и индуцированных переходов совместно позволяют описать результирующее равновесие, не зная ничего о деталях кинетики его установления. С задержкой почти на полстолетия эти коэффициенты привели к созданию лазеров, без которых уже невозможно представить себе современную технику и жизнь людей.
В твёрдых телах объединение энергетических уровней электронов составляющих их атомов становится энергетическими зонами Бриллюэна (для биологов достаточно популярно об этом см. [23], глава IV, параграфы 4 – 6, глава V, параграф 1). Для органических и биомолекул возникает подобие таких зон, отличающееся ничтожным объёмом по отношению к зонам Бриллюэна в твёрдых телах. Отражает это известный в химии метод молекулярных орбиталей. Надо отметить, что метод молекулярных орбиталей (как практический) применим только для относительно простых органических соединений. Однако в конечном счёте биохимические реакции являются электронными переходами на уровне зон-орбиталей. «Мгновенный» в единичном атоме переход электрона при излучении и поглощении света для биомолекул превращается в цепочки реакций и промежуточных веществ, отражающих перестройки электронных орбиталей. Примером такого могут служить цепочки цитохромов, участвующие во многих реакциях метаболизма. Свои относительно большие зоны-орбитали имеют молекулы РНК и ДНК. Детали их структуры зависят не только от индивидуальности этих молекул, но изменяются с участием водородных связей в зависимости от их белкового окружения, то есть от структуры и метаболизма хромосом в клетке.
Одной из основ образования новых таксономических градаций являются изменения ДНК, которые известны как мутации. В терминах рассказанного выше мутация является электронным переходом в молекуле ДНК, выраженным химическими реакциями, их продуктами и результирующими перестройками этой молекулы. Поэтому установление равновесия, соответствующего мутации, может быть описано с помощью феноменологических коэффициентов спонтанных и индуцированных переходов в молекуле ДНК, минуя описание промежуточных химических реакций и их продуктов. Однако до работы [2] (глава IV, параграф 6 «Спонтанные и индуцированные процессы как составляющие мутагенеза») никто не понял и даже не упомянул мельком роли в образовании мутаций и новых таксономических градаций жизни спонтанных и индуцированных переходов в том смысле, в котором их ввел А. Эйнштейн для излучения и поглощения света. Кстати, сами слова – спонтанные мутации и индуцированные химические реакции – в генетике используются, но физический смысл их при этом утерян.
Для использования метода индуцированных и спонтанных переходов [2], [8] в задаче о развитии жизни (то есть о возникновении её новых таксономических градаций и изменений внутри существующих) достаточно знать из наблюдений и экспериментов, что существуют воспроизводимые реальные состояния молекулы ДНК, или хромосом в клетке, или даже только самих клеток, которые можно обозначить любым образом. О том, что это именно состояния, соответствующие возможным путям развития жизни, свидетельством является наличие между ними хотя бы сколь угодно редких спонтанных (случайных) переходов. Они не обязательно должны проявлять себя как начало какого-либо осуществлённого развития. Относительное энергетическое положение этих состояний не имеет значения. Феноменологически существуют между ними спонтанные переходы с большой или малой вероятностью – значит возможны такие условия, при которых включатся индуцированные процессы со своими феноменологическими характеристиками. В зависимости от величины констант, соответствующих спонтанным и индуцированным переходам, время и детали установления равновесия будут разными, но в некоторых внешних и внутренних условиях оно будет достигнуто. В открытой системе, каковой является жизнь, оно будет преимущественно динамическим. Хотя существуют споры бактерий как предел «неживого» статического равновесия молекулы ДНК. Переход её в форму споры так же содержит спонтанную и индуцированную составляющую.
Свершившаяся мутация есть результат установления нового равновесия. Хорошо подтверждает сказанное выше известный в генетике факт, что лучевые и химические мутагены вызывают мутации, которые наблюдаются как редкие спонтанные. В [2] (глава IV, параграф 6) приведено перечисление п.п. a – m известных фактов генетики, которые дополняют сказанное выше. Этот список, несомненно, можно улучшить и продолжить. Надеюсь, что [2] и эта статья побудят генетиков посмотреть на известные им факты как основания для практического применения введенного в [2] способа для анализа истории и образования новых таксономических градаций жизни на основе спонтанных и индуцированных процессов. Это даст возможность улучшить таксономическую систематику, так как существующая не удовлетворяет активно работающих биологов (в том числе систематиков). Примеры есть в [20], [21].
Описание мутаций и образования новых таксономических градаций жизни на основе феноменологических коэффициентов спонтанных и индуцированных переходов устраняет ещё одну трудность в генетике. Дело в том, что для описания процессов с участием ДНК в генетике используется как язык хромосом, так и язык двойной спирали ДНК. В учебниках генетики (даже последних лет, например, [24]) отмечается, что надёжный мост между ними пока не построен. Состояния, между которыми происходит переход, приводящий к новым равновесиям (мутациям) и последующему синтезу информации (самоорганизации) на их основе как элементов системы, могут быть связаны как с изменениями в двойной спирали ДНК, так и с перестройками хромосом с участием ДНК.
Простые примеры того, что язык хромосом должен сильно отличаться от языка ДНК. Кольцевая ДНК кишечной палочки имеет длину 1,4 мм, а заключена в клетку, имеющую форму палочки с длиной всего 2 мкм и диаметром 1 мкм. Ядерная ДНК эукариотических клеток имеет длину 2 м, а в стадии интерфазы заключена в ядре клетки, у которого диаметр меньше 10 мкм. Если клетка находится в стадии митоза, то её ДНК как клубок становится ещё меньше. В «упаковке» ДНК в клетке прокариот и в ядрах клеток эукариот участвуют катионы (двухвалентных металлов), а также полиамиды или белки. Формируют силы взаимодействия в этом случае электростатические силы между отрицательно заряженными фосфатными группами и нейтрализация зарядов положительными ионами металлов, полиаминами, или остатками белков. Поэтому упаковка ДНК в хромосомах связана с существенными изменениями её энергии. Учёт этого в рассуждениях о митозе и мейозе отсутствует. Чем отличаются X и Y-хромосомы с точки зрения упаковки в них ДНК и энергии для этого? Пожалуй, найти ответ на этот вопрос даже не пытались. Это только одна из энергетических проблем, например, для гамет.
О водородных связях в химии и биохимии и их широком участии в живых системах известно много. Но не установлены их общие принципы в комплексе свойств РНК и ДНК как молекул живых систем. При этом важно, что воду, роль которой в живых системах никто не преуменьшает, отличает от других растворителей примат водородных связей.
Приведу ещё пример такого рода.
Количество ДНК (двойной спирали) в хромосомах диплоидных соматических клеток для одного и того же биологического вида постоянно с хорошей точностью. Рентгеноструктурный анализ непосредственно в головках сперматозоидов, которые в отличие от соматических клеток имеют гаплоидные хромосомы, показывает, что ДНК в них имеет форму двойной спирали [25]. Одновременно непонятный пока достоверный факт – в сперматозоидах количество ДНК меньше в два раза, чем в соматических клетках [25]. Например, у кур в соматических клетках оно колеблется в разных органах в пределах от 2,7 до 2,3 пикограмм (10 –12 г ) на ядро клетки, а в сперматозоидах у них же составляет 1,3, у карпа это 3.3 – 3,5 и в сперматозоидах 1,6, у быка 6,3 – 6,4 и соответственно 2,8. Существуют полные организмы в виде, например, трутней у пчёл, крылатых особей-самцов в муравейниках, у которых в соматических клетках гаплоидный набор хромосом (а не диплоидный как у дословно их собратьев).
Гаплоидные хромосомы достоверно являются важнейшим для развития жизни феноменологическим состоянием (в том числе энергетическим для ДНК и её окружения), но в терминах последовательностей химических реакций и их кинетики связь гаплоидности непосредственно с нитями ДНК пока не имеет достаточного описания. Если систематизировать имеющиеся экспериментальные данные и собирать новые в терминах феноменологических постоянных спонтанных и индуцированных переходов, то во многих задачах такой пробел не будет существенным. Это ещё раз подчёркивает важность введения в [2] в генетику феноменологических коэффициентов спонтанных и индуцированных процессов.
Иллюстративно часть переходов в молекуле ДНК можно отобразить схемой на рис.6. На ней в овале внизу надпись «Двойная спираль» обозначает основное состояние молекулы ДНК. Слово – основное – здесь имеет смысл термина атомных энергетических уровней в физике. Оно является статически равновесным. Достигается оно у прокариот при переходе клетки в форму неживой молекулы – споры. У эукариот основное состояние соответствует динамическому равновесию. Оно реализуется при стационарном состоянии ДНК в динамики метаболизма живого организма. Противопоставляются основному состоянию в терминах возбуждения в атомной физике, например, динамические состояния при репликации в процессе деления клетки.
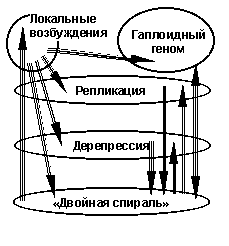 Рис.6
Рис.6
Состояние ДНК в гаплоидных хромосомах, (например, в сперматозоидах) существенно отличается от основного состояния, обозначенного на рис.6 «Двойная спираль» (теперь в смысле динамического равновесия), тем, что в составе живого одноклеточного организма, каковым является сперматозоид, ДНК теряет почему-то способность к локальным возбуждениям, необходимым для размножения этого одноклеточного организма. Поэтому естественно присвоить энергетическому состоянию ДНК в гаплоидных хромосомах название самого высокого уровня возбуждения. Энергетический механизм такого состояния не обязательно совпадает с механизмом локальных возбуждений.
Достоверно существуют организмы, у которых соматические клетки гаплоидные. Выше приводились примеры таких организмов в виде трутней у пчёл и крылатых муравьев-самцов. Это значит, что белковое окружении ДНК в хромосомах у таких особей другое, чем у одноклеточных автономных организмов – сперматозоидов. Оно снимает блокировку энергетических возбуждений ДНК, необходимых для процессов размножения клеток. Снимает её и проникновение в женскую яйцеклетку гаплоидных мужских хромосом. Такое известно как половое размножение. В генетике пока нет даже попыток объяснить эти детали на единой основе, полагающейся для столь фундаментальных процессов.
Феноменологическая расшифровка геномов разных видов живых организмов сегодня доведена до уровня технологического «заводского» процесса. Количество и разнообразие расшифрованных геномов растут с большой скоростью. В терминах введенной в [2] и пояснённой здесь обобщающей теории видообразования расширяется возможность систематизировать и практически использовать феноменологические константы спонтанных и индуцированных процессов в ДНК при анализе образования таксономических градаций жизни. Нужно доброжелательное участие генетиков для того, чтобы на основе [2] – [8] превратить иллюстративную схему на рис.6 в реальные феноменологические состояния ДНК с участием белков, формирующих хромосомы.
6. Экспериментальные подтверждения в биологии закона иерархического синтеза действия-энтропии-информации
В статье [3], в которой первично введен закон иерархического синтеза информации, на стр. 112 в качестве примера его применения предсказано, цитирую: «количества информации, отличающие простейшие бактерии друг от друга или от простейших многоклеточных организмов несопоставимо велики по отношению к количеству информации ответственному за мозг». Расшифровка генома человека [26], [27] дала результат, что около трети от всех человеческих генов совпадает с микробными, как и должно было быть при экспоненциальном падении количеств информации внутри последовательных ступеней иерархии, предусмотренном этим законом. Однако для биологов и генетиков это оказалось в результатах [26], [27] одной из больших сенсаций, так как в силу традиционного подхода (но ошибочного) все были убеждены, что генетическая информация собственно об отличиях человека от предыдущих таксономических градаций должна быть больше, чем об индивидуальных отличиях любого из его эволюционных предшественников. Не так много в истории науки новых законов, предсказания на основе которых получали бы столь весомое и однозначное подтверждение.
Ещё одно экспериментальное подтверждение решающей роли в эволюции жизни закона иерархического синтеза действия-энтропии-информации опять пришло из расшифровок геномов [28]. В этом случае генома простого многоклеточного вида жизни – актинии Nematostella. В её ключевом положение по отношению ко всем последующим видам жизни можно убедиться, например, по книге палеонтолога Стефана Гулда [29]. Актиния Nematostella относится к первичным представителям многоклеточных животных, которых систематики называют Eumetazoa.
Рассмотрю в чём и как экспериментальные исследования, отражённые в [28], подтверждают результаты и предсказания в [3] и [2] – [8].
Согласно схеме рис. 5 решающий шаг перехода от неживой природы к жизни связан с иерархически новым источником случайностей в виде энергетического равноправия многих разных комбинаций расположения кодонов в составе ДНК. Главным условием, ограничивающим эти случайности и синтез информации на их основе, является принцип структурной комплементарности. В результатах, приведенных в [28], отмечается, что около 80% генов человека совпадают с генами актинии и что они примерно в таком же количестве содержатся у её предков. Этот результат исследования [28] показывает, что возникновение жизни во всём её последующем разнообразии было скачкообразным переходом на следующую ступень иерархии синтеза информации, который создал «одномоментно» возможность работы критериев 1 – 7 на рис.2 и этим основу для самоорганизации всех последующих таксономических градаций жизни – как и предсказано в [2] – [8], генетический анализ в [28] достоверно, количественно показал, что именно синтез информации положил начало существующим сегодня таксономическим градациям жизни. Причём во всём её разнообразии, так как эти около 80% относятся не только к генам, но и к их порядку расположения в хромосомах. Более того, это продолжается вниз к одноклеточным организмам, то есть до ступени иерархии возникновения предшественников Eumetazoa. Вверх такие аналоги имеются у существующих даже высших многоклеточных организмов. Опять-таки это убедительно подтверждает предсказание в [3] на основе закона иерархического синтеза действия-энтропии-информации, то есть его справедливость.
Ещё одна немаловажная особенность этих совпадений в том, что они проявляются и для форм жизни, которые таксономически не относят к животным, то есть которые стали самостоятельными (иногда тупиковыми) ветвями эволюции жизни. Это означает, что контроль условий принципа структурной комплементарности над случайностями перестановок кодонов в ДНК оказался настолько жёстким, что признаки элементов, на основе которых произошёл этот синтез информации, на протяжении то ли около 3,5 миллиарда лет от возникновения прокариот, то ли 0,7 миллиарда лет от эукариот не смогли измениться, а стали основой локальных синтезов информации согласно критериям 1 – 6 на рис.2.
Были ли эукариоты скачком в синтезе информации по отношению к прокариотам или их ранг ниже? – Без генетических исследований ответить на этот вопрос нельзя, но симбиотическая гипотеза Л. Саган – Л. Маргелис, достоверность которой особых сомнений не вызывает, говорит в пользу первичности скачка для прокариот (об этой известной гипотезе см., например [2], глава II, параграф 7 «Способ питания прокариот – фагоцитоз – основа возникновения эукариотической клетки»).
Согласно результатам исследования [28], например, мухи и круглые черви в своём геноме имеют меньше аналогов генов предшественников актинии, чем человек – примерно 50% для мух и 40% для круглых червей. И это подтверждает первичность возникновения скачком особенностей элементов, на основе которых самоорганизация согласно критериям 1 – 7 рис. 2 универсально привела к существующим формам жизни на Земле. Кстати, этот результат не вписывается в «борьбу за существование», так как последовательности типа продолжения старого в новом в нём нет. Приведенный пример случайных больших потерь генетического материала у мух и червей показывает, что не отбраковка, а синтез информации, использующий источник случайностей в виде энергетически равноправных форм ДНК, является источником нового видообразования. С учётом результатов расшифровки генома актинии [28] количества генетической информации, ответственной за мозг человека (за его разум и сознание), ничтожно, как и написано в цитате из [3] выше.
Убедительно подтверждают справедливость новых принципов эволюции жизни и видообразования, введенных в работах [2] – [8], исследования микробных сообществ, отражённые [18], [20], [21] (см. также ссылки в [2] на предыдущие публикации Г.А. Заварзина). В [30] содержится краткий обзор представительного международного рабочего совещания в Новосибирске по проблеме «Происхождение и эволюция биосферы». Его цель была в выявлении наиболее значительных достоверных фактов в истории жизни на Земле (преимущественно относящихся к прокариотам, а также миру РНК до них и эукариотам после них). Практически все доклады там можно использовать как экспериментальное подтверждение нового подхода к возникновению и эволюции биосферы, изложенного в [2] – [8] и в этой статье. Но в силу причины, отмеченной здесь в примечании 5, для детального обсуждения экспериментальных фактов необходимо желание исследователей понять их с позиций закона иерархического синтеза действия-энтропии-информации. В этом есть трудности с их стороны. В [30] Г.А. Заварзин отмечает: «академики проявили гораздо больше скепсиса и стремления к пересмотру существующих концепций, чем их молодые коллеги, предпочитающие оставаться на более прочной основе традиционных подходов». Такое действительно наблюдается (даже в агрессивных формах).
7. «Апериодический кристалл» Э. Шредингера в терминах иерархического синтеза действия-энтропии-информации
В книге [1] содержится интуитивное предчувствие Эрвина Шрёдингера, что в основе жизни должен быть некий «апериодический кристалл». Оно получило детальное выражение, допускающее физическую и математическую интерпретацию, после введения в [3] закона иерархического синтеза действия-энтропии-информации. Поясню это на основе [2] глава VI, параграф 13 «Квазикристаллы как важнейшие структуры жизни».
Действие-энтропия-информация определено в шестимерном фазовом пространстве конфигурационных координат и импульсов (упрощено – скоростей). Наглядно это означает, что в природе от субатомного уровня и выше первичным является движение и положение как неразрывное целое. Обычные кристаллы и экстремумы свободной энергии в большинстве задач химии создают формы, которые описываются сечениями фазового пространства конфигурационным пространством под рациональными углами. В отличие от этого молекулы РНК и ДНК существуют в сечениях фазового пространства, когда угол этого сечение конфигурационным пространством иррациональный. С такими сечениями связано понятие квазикристаллов, симметрия которых не видна в трёхмерном пространстве (то есть на рисунках). Они существуют и в неживой природе, например, в металлах. Но там они дают однозначные результаты, а для РНК и ДНК в этих сечениях энергетические критерии порождают случайности многих пространственно разных последовательностей кодонов. Судя по всему, Э. Шредингер о квазикристаллах не знал (хотя в математике они появились раньше его книги [1], в 1903 г.). Но он назвал будущую молекулу ДНК «апериодический кристалл». Работа [2] глава VI, параграф 13 впервые ввела в задачу о РНК и ДНК понятие квазикристаллов и объяснение этим сочетания энергетических закономерностей и случайностей для молекул РНК и ДНК.
Закон иерархического синтеза действия-энтропии-информации даёт ответ – жизнь во Вселенной является множественным явлением и обязательно на основе углерода. Кремнийорганическая (и тем более на основе других элементов) жизнь во Вселенной невозможна, если принять, что химические элементы одинаковы во всей Вселенной. Что касается деталей путей её возникновения и первичных форм, то даже на Земле диапазон реализации жизни по температуре окружающей среды, давлениям и химическим условиям шире обиходно привычного. Пока неясно являются или нет повышенные температуры и экзотические условия вулканической деятельности обязательными для первичного зарождения жизни. Как отмечено в [2] глава V, параграф 8 «Самозарождение жизни в современных условиях», явно реализуемое сегодня быстрое образование новых видов в зоне вулканической деятельности на дне океанов («черные курильщики») говорит в пользу того, что первичная жизнь на Земле возникла в условиях высоких температур (по отношению к условиям для массово окружающих нас форм жизни).
8. Насколько «древовидно» развитие жизни?
В статьях Г.А. Заварзина, например [30], [31], справедливо выражается сомнение в сегодняшней абсолютизации эволюционной непрерывности дарвинизма в стиле указаний – самое главное найти, кто от кого произошёл. Это сомнение отражает реальность.
Джеймс Максвелл был более молодым современником Дарвина. Когда вышла книга Дарвина, ему было 27 лет. Он создатель электродинамики, поддержавший одним из первых работы по молекулярно-кинетической теории газов Людвига Больцмана. «Распределение Максвелла» было опубликовано им на год позже книги Дарвина. Максвелл, физик, почти на 100 лет раньше Э. Шредингера поставил один из двух главных вопросов его книги – о молекуле жизни. Он писал (см. [32], главаIV, параграф 3 «“Запас скрытых задатков” Максвелла»), что: «...Молекулярная физика ставит нас лицом к лицу с физиологическими проблемами. ... Микроскопический зародыш не есть лишь индивидуальное тело, он представитель, содержащий члены, собранные со всех ветвей широко раскинувшегося родословного дерева, и число этих членов вполне достаточно не только для того, чтобы передать наследственные особенности каждого органа тела и каждой привычки животного от рождения до смерти, но также и для того, чтобы дать возможность запасу скрытых задатков переходить в недеятельном состоянии от зародыша к зародышу, до тех пор пока особенности, представляемые им, не возродятся вновь в каком-нибудь отдалённом потомке» (выделено мною. А.Х.). Но как было показано в этой статье на основе [2] – [8], «запас скрытых задатков» не должен непосредственно содержаться в «молекуле жизни». Его создаёт новый иерархический уровень случайностей, реализованный в ДНК, в сочетании с принципом структурной комплементарности и законами квазикристаллов.
Объекты внутри каждой иерархической ступени синтеза действия-энтропии-информации (таксономических градаций жизни) существуют на основе свойств элементов предшествующих ступеней. Поэтому древовидная структура таксономической классификации жизни неизбежна. Но генетический код математически избыточен и неоптимален. Это дополняется в силу энергетического равноправия комбинаций кодонов в ДНК огромным количеством генетического материала, который не управляет белками – «молчит». В определённых условиях он может начать работать в том числе и совместно с работающими генами самых разных предыдущих таксономических градаций. Более того, может начать работать (и это нередкий случай) такая комбинация кодонов в ДНК, которая фактически не содержится в её «молчащей части», а возникла вновь в силу энергетического разрешения для такой случайности. В результате «дерево» таксономических градаций с «ветками» в любой своей части может дать «побеги», «наросты», которые содержат признаки, далеко отстоящие от данной «ветви» и места на ней или не встречавшиеся раньше. Если в схемах таксономического описании жизни их соединять с местами, где они раньше проявлялись, или могут внешне казаться изменениями в давно прошедшем, то таксономическое «дерево» окажется переплетенным «паутиной». Поэтому, например, неправильно говорить (как общепринято) человек «произошёл от обезьяны». Он является только одним из синтезов информации на «ветви» приматов, которая «запаутинена» со всем «деревом жизни» и внутри.
Причина того, что в природе существует жизнь, содержится в первичных законах физики, начинающихся от формирования «элементарных частиц» и атомно-молекулярной структуры неживых веществ. Что получится в результате – дерево, ветви или пересекающаяся паутина – зависит от подробностей внешних и внутренних условий в процессе данного иерархического синтеза информации. Решающий для возникновения жизни синтез информации произошёл на уровне прокариот. Следующая ступень эукариот и многоклеточных организмов основана на их свойствах как предыдущих элементов физической системы. Они остались решающими для всех последующих синтезов информации. Но это не запрещает локальных и глобальных «паутин».
«Паутина» – это напоминание о разбросе в виде разрывов непрерывности при реализации случайных процессов в разных условиях.
У основания ствола «запаутиненного дерева жизни» есть остановившиеся в развитии тупики, например, высокотемпературные бактериальные формы жизни. Они очень мало исследованы. Есть сомнения в полном единстве их ДНК и белков с состоявшимся «стволом». Но принцип максимума способности превращений выделяет основной «росток», который саморегулирует внешние условия. Последнее хорошо исследовано в работах Г.А. Заварзина, отражённых, например в [18], [20], [21].
В природе система всегда возникает и развивается вперёд во времени. Отличие жизни от привычных систем в науке и технике состоит в физико-химическом факте – исходные для жизни молекулы РНК и ДНК не имеют однозначности линейной компоновки составляющих их групп – кодонов. Поэтому в любой момент развития живой системы в этих молекулах возможны случайные перегруппировки составляющих. Трактовать это как влияние того, что «здесь и сейчас» на прошлое неправомерно. Более того, как правильно подчёркивается в работах Г.А. Заварзина, первичные формы жизни (обобщённо называемые бактериями) необратимо изменили саму планету Земля и этим ограничили случайности для РНК и ДНК на протяжении развития жизни в прошлом, настоящем и будущем.
Изменения в ДНК не становятся информацией сами по себе. Синтез информации на основе ДНК происходит только тогда, когда реализуется полная (сложнейшая!) цепочка многих иерархических синтезов информации, работающая от ДНК до клетки. Но и этого мало! Изменения клетки с участием изменения генетического кода в ДНК должны быть совместимы со всеми последующими иерархическими ступенями синтеза информации вплоть до ноосферы в целом. Это соответствует определению информации как физической переменной, особенности которого указаны на рис.1. Средства связи человека передают сигналы, а не информацию. Сигналы сами по себе не могут устранить неопределённость. Для человека, который не знает азбуки Морзе, её попискивание –только раздражающий шум. Знание её – это и есть «Книга сообщений» на рис.1. В природе предзаданной «Книги сообщений» нет, правая колонка на рис.1. Генетический код – это только сигналы для живой системы. Они становятся информаций после того, как на их основе реализована вся цепочка синтеза информации, для которой генетический код задал свойства исходных элементов живой системы. Поэтому нет в живом организме адресной (в смысле компьютеров) ячейки, в которой хранился бы какой-то феноменологический признак организма в целом или его функциональных органов. Данный ген в составе ДНК содержит в себе генетическую информацию постольку, поскольку с его участием может быть устойчиво воспроизведена (и тем запомнена) вся иерархическая цепочка синтеза информации, которая необходима от гена до макроскопического феноменологического признака организма. В наибольшей степени это проявляется в задачах иммунитета, где принципиальный прорыв в этом связан с именем И.Л. Черткова, который первым обратил внимание на стволовые клетки и провел их исследования вплоть до практических результатов. Работа аналогичных механизмов в эволюции жизни ещё только начинает исследоваться. Но уже ясно, что это введёт подробности в цепочки синтеза информации, ответственной за образование новых таксономических градаций.
9. Некоторые методологические замечания
Об абсолютной истине. Вопреки философам абсолютная истина существует (см. [2], [7]). Парадоксально ею являются утверждения и научные результаты, для которых известны границы, за пределами которых они ошибочны. В таком смысле дарвинизм является абсолютной истиной в пределах границ, которые исходно сформулировал сам Дарвин и которые забыты поколениями, наследовавшими ему.
О физическом принципе локальности взаимодействий в эволюции жизни. В основе физики уже столетия лежит принцип близкодействия Ньютона. Он подразумевает существование точки контакта тел. С появлением в физике понятия – поле (достаточно популярно для биологов о нём см. [22]) место контакта заменило понятие предела функции в точке. Однако ещё древнегреческие атомисты около 2500 лет назад ввели неделимый атом. Возник парадокс невозможности определить точку контакта тел, так как метка о ней будет запрещённой частью неделимого атома, а предельный переход невозможен из-за дискретности атомов. В результате в современной атомной физике ньютоновское близкодействие заменяет математическое требование калибровочной инвариантности. При случайных взаимодействиях близкодействие реализуется с участием осреднённых характеристик функций распределения и, в частности, понятия об энтропии. В этом случае близкодействие принимает форму, в которой участвуют флуктуации определяющих переменных. О введении действия-энтропии-информации в аксиоматику механики и квантовой механики см. [6].
В качестве математического средства при описании характеристик случайных процессов, в частности энтропии, используется комбинаторика. Природа содержит в составе реальных флуктуаций переменных следы того, что описывает математически комбинаторика. Флуктуации являются тем, что отличает статистическую реализацию ньютоновского близкодействия или его более сложного развития в других областях физики. Но природа не может и не должна реализовать математические приёмы комбинаторики как реальные пробы и ошибки.
Философы заменили строго сформулированные требования локальности взаимодействий у физиков терминологической эквилибристикой слов «здесь и сейчас» в отношении явлений природы и их описания. Это только запутывает дело. В частности, в [31] остро обсуждается вопрос, что дарвиновский естественный отбор противоречит рассуждениям философов о «здесь и сейчас». Как было введено в [2] и подчёркнуто выше в этой статье, дарвиновский естественный отбор является эмпирическим описанием частного случая принципа минимума производства энтропии по отношению к ней как действию-энтропии-информации. В силу объяснённого выше такая физическая основа гарантирует его соответствие требованию локальности явлений и описывающих их теорий. Недоразумения в связи с естественным отбором вызывает приписывание природе, якобы, «осуществление» как реальных проб и ошибок комбинаторных подсчётов (сугубо человеческого абстрактного математического приёма!). В полном соответствии с принципом локальности кинетика установления данного значения энтропии (действия-информации) для живых объектов однородна с аналогичным во всей природе (например, давление в газах, жидкостях и др.). Для жизни предыстория эволюции (развития) данной таксономической градации присутствует локально («здесь и сейчас» в терминах философов) потому, что элементами, на основе свойств которых происходит новый иерархический синтез информации (действия-энтропии), являются объекты предыдущего уровня иерархии.
Как отмечалось в этой статье, дарвиновский естественный отбор был сформулирован в десятилетие первичного введения в науку основных сегодня её терминов – энергия и энтропия. Парадоксальным образом он на полтора столетия остался в науке изолированным островом вне её физических основ. В [2] это устранено определением его места в науке как частного эмпирического варианта принципа минимума производства энтропии (действия-информации). Однако в научной литературе дарвиновский естественный отбор продолжает обсуждаться на основе обиходного понимания отбора как проб и ошибок, например, при подборе на заводе стержней и втулок к ним из заготовок с разбросом диаметров.
О целях в биологии и целесообразности. Конкретной цели возникновения феноменологических признаков организмов и их развития в природе нет и быть не может (см. [2] – [8]). Кажущаяся целесообразность в природе возникает потому, что принципы иерархического синтеза действия-энтропии-информации в равной степени первичны для неживой природы, жизни и для работы нервных систем, включая мозг человека. Поэтому природа разумна в той же степени, что и человек (кроме свободы воли в результате существования у человека двойной сигнальной системы, см. подробности в [33]). Потому полнота учёта законом иерархического синтеза действия-энтропии-информации взаимосвязей в природе между её объектами и процессами воспринимается человеком (то есть с учётом его свободы воли) как её целесообразность. Важно подчеркнуть, отмеченное в [2], что процветание человечества не предусмотрено законами природы как её цель. Это же (даже более жёстко) сказал Ю.А. Розанов в публикации [21], в дискуссии там: «...интересы человека и интересы биосферы находятся в полном противоречии, и задача биосферы человека уничтожить».
В истории жизни на Земле вымерло 99,9% когда-либо существовавших видов жизни. Вымрет и человечество, если не поймёт, что в его прошлом биохимически заложена взаимная корреляция разума и самоагрессивности. Может ли полученный с её участием разум и свобода воли человека защитить его от самоуничтожения? Ответа на этот вопрос пока нет! ***
Накануне 150-летия книги Чарлза Дарвина «Происхождение видов путём естественного отбора или сохранения избранных пород в борьбе за жизнь» нам всем остаётся ещё раз отдать должное величию его ступени вклада в прогресс науки и вспомнить его слова в этой книге: «...мои выводы были превратно истолкованы и утверждали, что я приписываю изменение видов исключительно естественному отбору. ... Я убеждён, что естественный отбор был главным, но не исключительным средством, вызывающим изменения. ...Но это не помогло. Великая сила извращения чужих мыслей».
Литература:
1. Шредингер Э. Что такое жизнь? М.: Атомиздат. 1972.
2. Хазен А.М. Разум природы и разум человека. М.: НТЦ Университетский. 2000. .
3. Хазен А.М. Происхождение и эволюция жизни и разума с точки зрения синтеза информации// Биофизика. Т. 37. В. 1. С. 105-122. 1992.
4. Хазен А.М. Принцип максимума производства энтропии и движущая сила прогрессивной биологической эволюции // Биофизика. Т. 38. N3. C. 531-551. 1993.
5. Хазен А.М. Особенности применения второго начала термодинамики к описанию работы мозга // Биофизика. Т. 36. №4. C. 714-724. 1991.
6. Хазен А.М. Введение меры информации в аксиоматическую базу механики. М.: РАУБ. 1998. (Первое издание М.: ПАИМС. 1996).
7. Хазен А.М. Первые принципы работы мозга, гарантирующие познаваемость природы. М. 2001.
8. Хазен А.М. Развитие на основе иерархического роста энтропии // Научный электронный журнал МГУ Физико-химическая кинетика в газовой динамике: http://www.chemphys.edu.ru/pdf/2005-10-06-006.pdf .
9. Полетаев И.А. Сигнал. М.: Советское радио. 1958. .
10. Голдман С. Теория информации. М.: Изд. Иностранной лит. 1957.
11. Фано Р. Передача информации. М.: Мир. 1965.
12. Хазен А.М. Законы природы и «справедливое общество». М.: УРСС. 1998.
13. Заварзин Г.А. Эволюция микробных сообществ. Доклад, прочитанный на теоретическом семинаре геологов и биологов «Происхождение живых систем», 15-20 августа 2003 г., Горный Алтай. Оригинал: http://www.bionet.nsc.ru/live.php?f=doclad&p=zavarzin .
14. Блюменфельд Л.А. Решаемые и нерешаемые проблемы биологической физики. М.: УРСС. 2002 г.
15. Блюменфельд Л.А. Отзыв о монографии А.М. Хазена «Разум природы и разум человека»//Биофизика. Т. 47. В. 2. С. 382. 2002 г.
16. Кастлер Г. Возникновение биологической организации. М.: Мир. 1967.
17. Блюменфельд Л.А. Информация, термодинамика и конструкция биологических систем // Соросовский образовательный журнал, №7, с. 88-92, 1996.
18. Заварзин Г.А. Составляет ли эволюция смысл биологии? // Вестник РАН, т. 76, №6, с. 522-543. 2006.
19. Спирин А.С. Рибонуклеиновые кислоты как центральное звено живой материи // Вестник РАН, т. 73, №2, с. 117-127. 2003.
20. Розанов А.Ю., Заварзин Г.А. Бактериальная палеонтология // Вестник РАН. 1997. №3.
21. Заварзин Г.А. Становление биосферы // Вестник РАН, т. 71, №11, с. 988-1001. 2001.
22. Хазен А.М. Поле волны, частицы и их модели. М.: Просвещение. 1979.
23. Хазен А.М. Современная электроника. М.: Просвещение. 1970.
24. Сингер М., Берг П. Гены и геномы. Т. 1. М.: Мир. 1998.
25. Волькенштейн М.В. Молекулы и жизнь. М.: Наука. 1965.
26. Nature. The Human Genome. V. 409. No. 6822. 15 February 2001.
27. Science. The Human Genome. V. 291. No. 5507. 16 February 2001.
28. Putman N., Srivastava M., Hellsten U., Dirks B., Chapman J., Salamon A., Terry A., Shapiro H., Lindquist E., Kapitonov V., Jurka J., Genikhovich G., Grigoriev I., Lucas S., Steele R., Finnerty J., Technau U., Martindale M., Rokhsar D. Sea Anemone Genome Reveals Ancestral Eumetazoan Gene Repertoire and Genomic Organization // Science. 6 July 2007. V. 317. P. 86-94.
29. Gould S. The Book of Life. W. Norton&Company. New York, London. 1993.
30. Заварзин Г.А. Эволюция биосферы с точки зрения геологов и биологов // Вестник РАН, т. 76, №2, с. 166-168, 2006.
31. Заварзин Г.А. Бытие и развитие: эволюция, сукцессия, хаэссеитас // Вестник РАН, т. 77, №4, с. 334-340, 2007.
32. Хазен А.М. О возможном и невозможном в науке, или где границы моделированимя интеллекта. М.: Наука. Главная редакция физико-математической литературы. 1988.
33. Хазен А.М. Функции разума и сознания в неживой и живой природе и их совместная эволюция // В процессе публикации. Препринт см. http://www.kirsoft.com.ru/intell/KSNews_112.htm .
Электронные адреса тех из книг списка литературы, которые есть в интернете, указаны на титульной странице сайта автора «Разум природы и разум человека» http://www.kirsoft.com.ru/intell . Дополнительные пояснения к статье есть в статьях на этом же сайте. Их можно выбрать из его «Содержания» по адресу http://www.kirsoft.com.ru/intell/KSNews_idx1.htm и прочесть. .
Данная статья является препринтом статьи, отправленной для публикации в научный печатный журнал.





